
摘要
微生物在稻田土壤中介导砷的解毒作用,决定了砷在土壤中的行为和对水稻的可利用性,但对低砷稻田中功能基因的结构和丰度以及驱动力知之甚少。为了探究砷解毒功能基因的模式,从中国四个气候带的39个稻田中收集了429份土壤样品,其砷含量范围为9.76~19.74 mg kg-1。利用GeoChip分析参与砷解毒的功能基因,共检测到三个砷解毒基因:aoxB、arxA和arsM。功能基因的多样性和丰度在不同的采样点之间都有明显的差异,并沿砷梯度下降。砷解毒基因由细菌、古细菌和真核生物携带。冗余分析(RDA)表明,土壤样品根据它们所在的气候带和大陆尺度的砷梯度进行分组聚类。土壤pH值、年平均温度(AAT)、土壤砷含量、年平均降水量(AAP)和CEC是塑造功能结构的最重要因素。结构方程模型(SEM)显示AAT、pH和土壤砷含量直接影响砷解毒基因丰度。这些发现提供了稻田土壤中参与砷解毒的微生物群落的整体情况,并揭示了气候因素在大空间尺度上塑造功能基因的重要性。
研究背景
砷(As)是一种普遍存在的有毒环境物质,通过地球化学过程和人类活动在地球圈之间和内部循环,通过食物链危害人类健康。稻田中的砷污染已成为世界范围内一个严重的健康问题,尤其是在稻米消费量高的亚洲和拉丁美洲。由于水稻土壤处于淹水厌氧条件,水稻比其他谷类作物(如小麦和大麦)更有效地吸收砷。最近的一项研究显示,中国稻米中的总砷浓度在0.01-0.186 mg kg-1之间,其中无机砷和二甲基砷酸分别占69%和31%。
微生物是砷地球化学循环的关键驱动力,催化砷的氧化还原和甲基化。As(V)还原的微生物途径是由胞内arsC还原酶和周质arrA还原酶介导的。微生物对As(III)的氧化是由aroA,aoxB和arxA编码的As(III)氧化酶介导的。As(III)被认为比As(V)毒性更大,因为无机As(III)可以与蛋白质中小分子硫醇和半胱氨酸残基的巯基反应,抑制生物体中重要的生化过程。此外,As(III)可被微生物甲基化生成有机砷。这一过程是由arsM基因编码的S-腺苷甲硫氨酸甲基转移酶催化的。虽然As(III)的甲基化产物比其本身毒性更大,但arsM催化的生物甲基化的最终产物是挥发性三甲基砷(TMA(III)),这最终导致介质中砷的减少。因此,As(III)的氧化和甲基化在三价砷的解毒中具有潜在的重要意义,这为砷污染土壤的生物修复提供了应用前景。
As(III)氧化微生物广泛存在于自然界,包括异养微生物和化学/光合自养微生物。根据基于PCR的方法研究,水稻土中砷氧化细菌的主要类群是典型的根际微生物,即变形杆菌中的根瘤菌和伯克霍尔德菌。厌氧微生物是厌氧环境中砷甲基化的主要驱动因素。在水稻土中,含有arsM的细菌主要属于变形杆菌属(Proteobacteria)、双生孢子目(Gemmatmonadales)和藻类(Firmiccus),具有较高的系统发育多样性。到目前为止,对砷生物转化微生物的描述主要集中在细菌上。用高通量技术揭示砷解毒微生物群落的多样性,对于更好地理解微生物介导的砷解毒具有重要意义。
微生物砷解毒基因的多样性和丰度共同决定了砷生物转化的方向和速率,从而影响土壤中砷的含量和形态。因此,了解微生物砷解毒功能基因的影响因素,对于进一步研究土壤砷污染的生物修复具有重要意义。到目前为止,关于低砷水稻土中砷解毒基因在大空间尺度上的分布及其可能的环境决定因素的信息还很缺乏。本研究利用基因芯片技术,对我国13个水稻主产区的39块稻田的砷解毒微生物群落进行了分析,以评估低砷水稻土中砷解毒相关基因的分布、多样性和丰度,揭示水稻土中砷解毒微生物群落的组成,及确定在大空间尺度上形成微生物砷解毒功能基因的重要环境驱动因素。
结果
01
土壤中总砷浓度的地理差异
通过对中国主要水稻种植区的429个水稻土样本进行采样&#xff0c;观察到采样点之间土壤总砷含量存在显著差异&#xff08;p<0.05&#xff09;&#xff0c;范围从9.76±1.43 mg kg-1到19.74±3.17mg kg-1&#xff08;图1&#xff09;。位于亚热带的ZX、CT和HY的土壤砷含量相对较低&#xff0c;平均为9.76&#xff5e;14.41 mg kg-1&#xff1b;中温带、暖温带和亚热带的HL、YY、SY、FQ和JO的砷含量平均为15.66&#xff5e;17.46 mg kg-1&#xff1b;中温带、亚热带和热带地区的QZ、CC、HK、LA和QX的砷含量较高&#xff0c;平均浓度为18.77&#xff5e;19.74 mg kg-1。根据中国土壤环境质量国家标准&#xff08;GB15618-2018&#xff09;&#xff0c;研究中所有土壤样品的砷含量均在风险筛选值的限度内&#xff0c;代表对农产品安全的低风险。也测定了其他土壤地球化学属性&#xff0c;包括pH、TN、TDN、NH4&#43;-N、NO3--N、DON、TP、AP、TK、AK、OM、DOC和CEC&#xff08;表S2&#xff09;。通过对土壤地球化学属性进行DCA分析&#xff0c;本研究发现稻田土壤样品在地理上是分组的&#xff0c;表明13个地区的地球化学特征存在显著差异&#xff08;图S2&#xff09;。
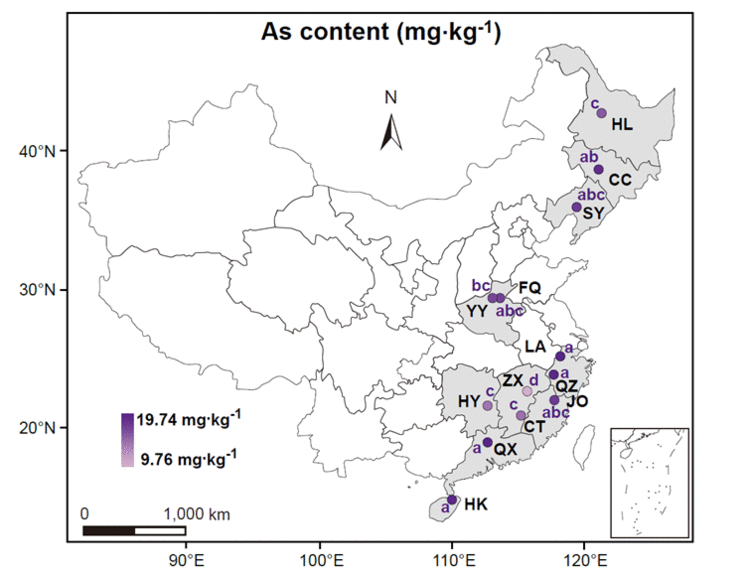
图1. 中国主要13个水稻种植区39个稻田的土壤砷浓度。
02
稻田土壤中砷解毒基因的多样性和丰度
在429份稻田土壤样品中检测到参与砷解毒的3个基因家族&#xff1a;aoxB、arsM和arxA。每两个土壤样品之间重叠基因的百分比从67.41%到98.39%不等&#xff0c;表明整个稻田土壤的基因组成具有高度相似性&#xff08;图2a&#xff09;。根据每个站点中独特基因的百分比&#xff0c;观察到沿着土壤砷梯度的独特基因比例在增加。总体而言&#xff0c;砷解毒基因的丰富度、Shannon-Wiener和Simpson指数在不同地点之间显示相当大的差异&#xff08;图2b和图S3&#xff09;。Spearman相关分析发现&#xff0c;稻田土壤砷解毒基因多样性与土壤砷含量呈显著负相关&#xff08;表1&#xff09;。水稻土中砷解毒基因的总体丰度&#xff08;GeoChip信号强度&#xff09;在41万~51万之间&#xff0c;其中72%为aoxB基因&#xff08;与As(III)氧化有关&#xff09;&#xff0c;17%为arsM基因&#xff08;与砷甲基化有关&#xff09;&#xff0c;其余11%为arxA基因&#xff08;与As(III)厌氧氧化有关&#xff09;&#xff08;图2c&#xff09;。在不同地区的土壤中发现属于aoxB和arsM家族的基因丰度存在明显差异。然而&#xff0c;关于arxA基因&#xff0c;样本之间没有明显差异。Spearman相关分析表明&#xff0c;arsM的丰度与砷浓度呈负相关。
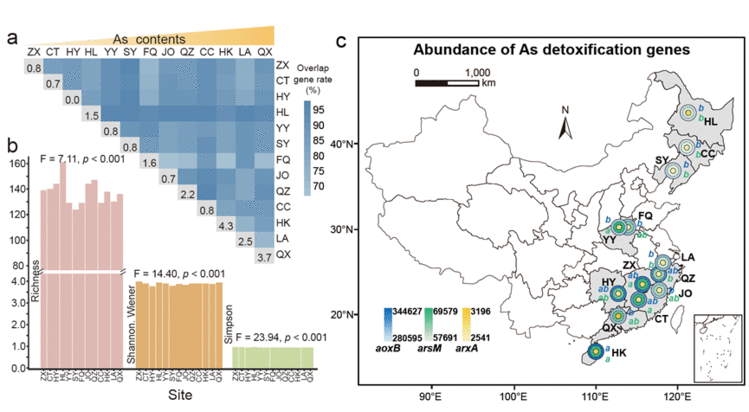
图2. 水稻土壤中砷解毒基因的多样性和丰度。(a) 每两个土壤样品之间重叠基因比率的热图。(b) 水稻土中砷解毒基因的丰度、Shannon-Wiener指数和Simpson指数。(c) 土壤中参与砷解毒的三个基因aoxB、arsM和arxA的丰度。
表1. 土壤中砷解毒基因与环境因子的Spearman相关系数。
03
砷解毒基因微生物群落组成
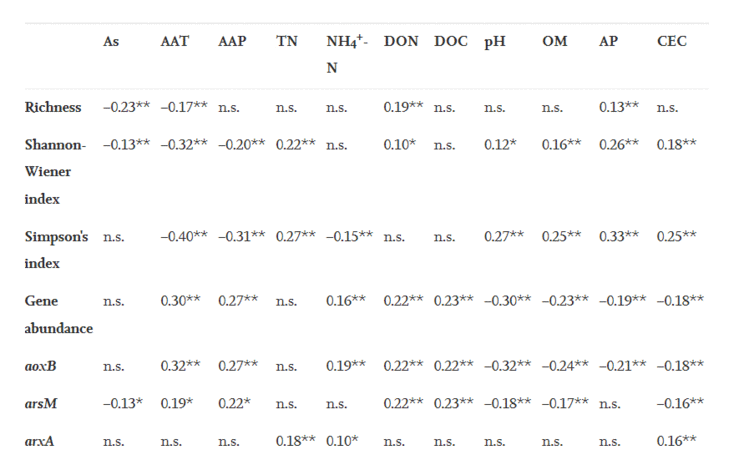
土壤细菌、古菌和真核生物携带的砷解毒基因的信号强度如图3a和3b所示。总体而言&#xff0c;细菌携带的砷解毒基因的信号强度在不同地点之间存在明显的差异&#xff0c;并沿着土壤砷梯度下降。所有细菌均隶属于17个门&#xff0c;其中Proteobacteria是所有样品中最主要的细菌分类群&#xff0c;占细菌携带的总基因丰度的55.79~59.09%&#xff0c;其次是Actinobacteria、Chloroflexi和Firmicutes。通过进行Spearman相关性分析&#xff0c;观察到砷含量与Acidobacteria、Actinobacteria、Firmicutes、Gemmatimonadetes、Nitrospirae、Planctomycete和Spirochaetes呈显著负相关。砷氧化细菌的主要类别为Proteobacteria和Actinobacteria&#xff0c;分别占细菌携带的总基因丰度的40.63~50.36%和26.27~35.43%&#xff08;图3c&#xff09;。Delta-proteobacteria对砷氧化Proteobacteria的贡献最大&#xff0c;其次是gamma-proteobacteria和beta-proteobacteria&#xff08;图S4&#xff09;。本研究还研究了所有样品中砷甲基化细菌的细菌群落组成&#xff08;图3d&#xff09;。结果表明&#xff0c;Proteobacteria占砷甲基化细菌的大部分&#xff0c;其次是Chloroflexi。Gamma-proteobacteria对砷甲基化的贡献最大&#xff0c;其次是alpha-proteobacteria和beta-proteobacteria。
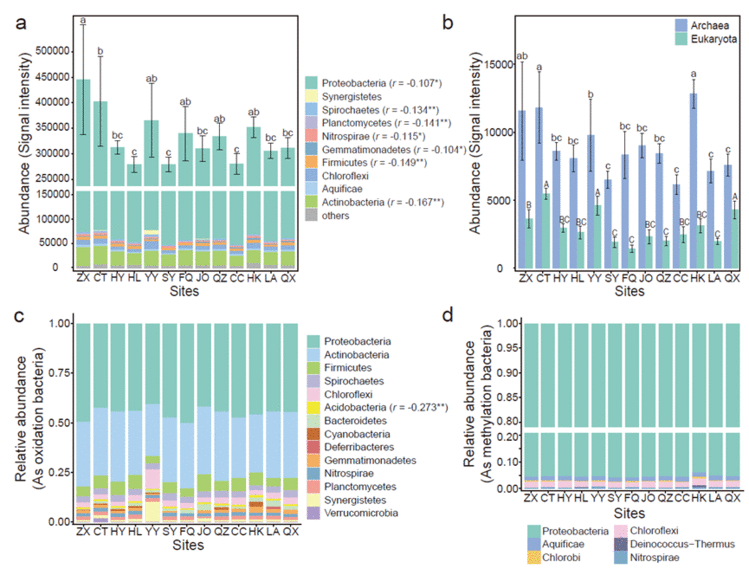
图3. 水稻土壤中砷解毒基因微生物群落组成。(a&#xff0c;b) 细菌、古细菌和真核生物携带的砷解毒基因丰度。在门水平上参与砷氧化(c)和甲基化(d)的基因的细菌群落组成。
04
参与砷解毒的微生物功能结构和基因丰度与环境变量的联系
进行冗余分析&#xff08;RDA&#xff09;以辨别参与砷解毒的微生物功能结构与环境变量之间可能的联系&#xff08;图4&#xff09;。RDA第一个轴与As&#xff08;土壤砷含量&#xff09;、pH和NO3--N呈正相关&#xff0c;但与TN、DOC和TK呈负相关。第二个轴与AAT&#xff08;年平均温度&#xff09;、AAP&#xff08;年平均降水量&#xff09;和NH4&#43;-N呈正相关&#xff0c;但与CEC、TP和OM呈负相关。土壤样品根据它们所在的气候带进行分组&#xff0c;显示出与分层聚类观察到的相似模式&#xff08;图4a&#xff09;&#xff0c;样品也根据土壤砷梯度分布&#xff08;图4b&#xff09;。总之&#xff0c;在大陆尺度上&#xff0c;参与砷解毒的微生物功能结构的变化与pH、AAT、As、AAP和CEC密切相关。SEM用于测试重要环境预测因子与砷解毒基因丰度之间潜在的直接和间接关系&#xff08;图5&#xff09;。结果表明&#xff0c;AAT是直接影响砷解毒基因丰度的最明显参数&#xff0c;其次是土壤pH和As含量。AAP通过调节土壤pH和砷含量间接影响基因丰度。根据标准化的总效应&#xff0c;气候因素对基因丰度有积极影响&#xff0c;土壤pH值和砷含量对基因丰度有消极影响。本研究进一步对重要环境预测因子和aoxB、arxA和arsM基因分别进行了SEM分析&#xff08;图S5&#xff09;。结果表明&#xff0c;与As(III)厌氧氧化相关的aoxB基因丰度直接受AAT、pH和As的影响。As(III)氧化基因arxA丰度仅受土壤pH直接影响&#xff0c;而砷甲基化基因arsM丰度仅受As直接影响。
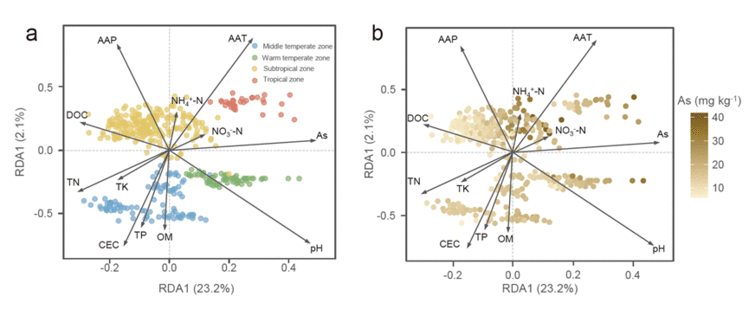
图4. 砷解毒微生物群落与环境变量的冗余分析。土壤样品分别按气候带(a)和土壤砷含量(b)进行分组。
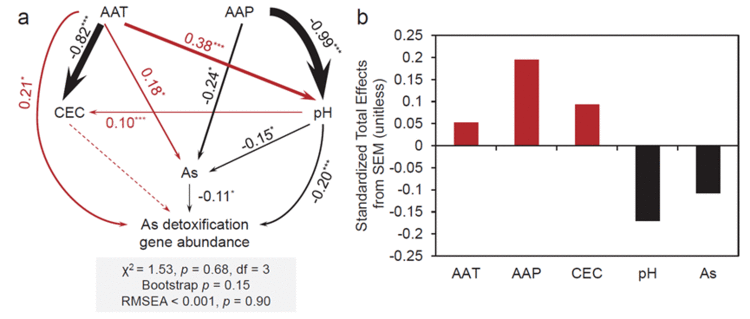
图5. 砷解毒基因丰度与重要环境变量因果关系的结构方程模型(a)以及标准化总效应(b)。
结论
本研究沿大空间尺度内稻田中的砷梯度分析了微生物砷解毒功能基因&#xff0c;包括aoxB、arxA和arsM。土壤微生物砷解毒基因的多样性和丰度在各采样点之间有明显的差异。与古细菌和真核生物相比&#xff0c;细菌是稻田土壤中含有砷解毒功能基因的主要群体。参与砷解毒的微生物功能结构受pH、AAT、As、AAP和CEC的影响。AAT直接对砷解毒基因丰度产生正向影响&#xff0c;而土壤pH值和砷含量对基因丰度产生负向影响。本研究不仅对砷解毒微生物群落的理解具有重要意义&#xff0c;而且还提供了潜在的信息&#xff0c;并有助于设计适当的策略&#xff0c;通过调节影响相关微生物的环境因素来刺激微生物进行场地砷的生物修复。
Reference

参考文献
Chi Zhang, Xian Xiao, Yuan Zhao, Jizhong Zhou, Bo Sun, Yuting Liang. Patterns of microbial arsenic detoxification genes in low-arsenic continental paddy soils. ENVIRONMENTAL RESEARCH, 2021. DOI:10.1016/j.envres.2021.111584
BACKPACK

作者简介
【第一作者】

张池&#xff1a;常州大学硕士研究生&#xff0c;主要关注逆境下水稻根际微生物的群落特征、相关功能基因及其对根系微生物组的调控过程。目前以第一作者发表了1篇ENVIRONMENTAL RESEARCH。
【通讯作者】

肖娴&#xff1a;女&#xff0c;博士&#xff0c;讲师。博士毕业于中国科学院南京土壤研究所&#xff0c;硕士及本科毕业于常州大学。2018年7月至今工作于常州大学环境与安全工程学院&#xff0c;主要从事土壤微生物生态与物质循环和土壤微生物修复技术等方面的研究。发表SCI论文16篇&#xff0c;中文核心论文5篇。主持国家自然科学基金青年科学基金项目1项&#xff0c;江苏省高等学校自然科学研究面上项目1项。
【通讯作者】

梁玉婷&#xff1a;中科院南京土壤研究所研究员&#xff0c;博士生导师。清华大学学士、博士&#xff0c;University of Oklahoma访问学者、博士后。从事土壤微生物学领域的研究&#xff0c;近年来在农田土壤微生物组功能与稳定机制方面取得了较为系统的研究成果,揭示了我国不同气候带典型农田土壤中微生物群落结构特征及碳氮耦合转化、地力提升功能的演变规律等。发表SCI 60余篇&#xff0c;中文核心10余篇。承担国家自然科学优秀青年基金、江苏省杰出青年科学基金等&#xff0c;中科院青促会优秀会员&#xff0c;担任《土壤学报》、Soil Ecology Letters等编委&#xff0c;中国土壤学会土壤质量标准化委员会主任、江苏省土壤学会学术工作委员会主任。

END
编辑 | 倪浩为
审核 | 梁玉婷
猜你喜欢
10000&#43;&#xff1a;菌群分析 宝宝与猫狗 梅毒狂想曲 提DNA发Nature Cell专刊 肠道指挥大脑
系列教程&#xff1a;微生物组入门 Biostar 微生物组 宏基因组
专业技能&#xff1a;学术图表 高分文章 生信宝典 不可或缺的人
一文读懂&#xff1a;宏基因组 寄生虫益处 进化树
必备技能&#xff1a;提问 搜索 Endnote
文献阅读 热心肠 SemanticScholar Geenmedical
扩增子分析&#xff1a;图表解读 分析流程 统计绘图
16S功能预测 PICRUSt FAPROTAX Bugbase Tax4Fun
在线工具&#xff1a;16S预测培养基 生信绘图
科研经验&#xff1a;云笔记 云协作 公众号
编程模板: Shell R Perl
生物科普: 肠道细菌 人体上的生命 生命大跃进 细胞暗战 人体奥秘
写在后面
为鼓励读者交流、快速解决科研困难&#xff0c;我们建立了“宏基因组”专业讨论群&#xff0c;目前己有国内外5000&#43; 一线科研人员加入。参与讨论&#xff0c;获得专业解答&#xff0c;欢迎分享此文至朋友圈&#xff0c;并扫码加主编好友带你入群&#xff0c;务必备注“姓名-单位-研究方向-职称/年级”。PI请明示身份&#xff0c;另有海内外微生物相关PI群供大佬合作交流。技术问题寻求帮助&#xff0c;首先阅读《如何优雅的提问》学习解决问题思路&#xff0c;仍未解决群内讨论&#xff0c;问题不私聊&#xff0c;帮助同行。

学习16S扩增子、宏基因组科研思路和分析实战&#xff0c;关注“宏基因组”
点击阅读原文





 京公网安备 11010802041100号
京公网安备 11010802041100号